Sejarah perkembangan pengetahuan sel
Pada tahun 1665, seorang Inggris,
Robert Hooke (1635-1703), mengamati sayatan tipis gabus botol dengan mikroskop
yang amat sederhana, yang dibuatnya sendiri. Yang terlihat olehnya adalah
struktur yang terdiri dari ruang kecil-kecil yang dinamakannya sel (cellula,
ruang kecil).
Nehemiah Grew (1641-1712)
menuliskan deskripsi pertamanya tentang jaringan tumbuhan pad a tahun 1671.
Pada tahun 1880, Hanstein menggunakan istilah protoplast bagi satuan
protoplasma dalam sel.
Pada tahun 1831, Robert Brown
menemukan nukleus dalam epidermis suatu anggrek. Hugo von Mohl melihat
perbedaan antara protoplasma dan cairan sel pada tahun 1846, dan pada tahun
1862 Kolliker memperkenalkan istilah sitoplasma. Sejak akhir abad 19 dan
selama abad ke-20 penelitian sel berkembang amat pesat sehingga membentuk ilmu
tentang sel atau sitologi.
Berdasarkan pada keberadaan nukleus
atau inti sel, dikenal dua kelompok organisme yakni prokariot yang tak memiliki
inti dan eukariot yang memiliki inti. Kelompok prokariot mencakup bakteri dan
ganggang biru. Organisme lain semuanya merupakan eukariot. Pada umumnya sel
mengandung sebuah inti saja, namun ternyata ada pula sel yang memiliki ban yak
inti, misalnya pada sel lateks (Iatisifer). Sel berinti-banyak sering dianggap
terdiri dari kumpulan inti yang masing-masing memiliki protoplas namun tak
berdinding. SeJ seperti itu, yang disebut senosit (coenocyte),menarik perhatian
dalam kajian fiIogenetik dan ontogenetik.
Ada dua macam teori yang berkaitan
dengan hubungan antara organisme secara keseluruhan dengan sel tunggal. Menurut
teori set yang dikembangkan pada pertengahan abad ke-19, organisme
terdiri atas kumpulan sejumlah besar sel yang masing-masing berperan dalam
menentukan sifat organisme yang bersangkutan. Teori lain adalah teori
o-rganismal yang tidak menonjolkan tiap-tiap sel, melainkan menekankan
kesatuan protoplasma seluruh organisme. Menurut teori ini, organisme-lah
sebagai kesatuan, sangat menentukan sifat sel pembentuknya.
Kedua teori tersebut mempunyai
kepentingan sendiri dalam pemahaman struktur tumbuhan. Banyak segi ontogeni
seperti proses pembelahan sel, asal mula unsur pembuluh kayu, dan perkembangan
idioblas diterangkan dengan mengingat teori seI. Namun, spesialisasi dari
macam-macam sel dan jaringan hanya dapat dijelaskan berdasarkan teori
organismal yang meninjau organisme sebagai suatu kesatuan.
2.3. Ciri
Dasar Struktur Sel
Sel
merupakan unit strukturan dan fungsional organisme hidup. Organisme terkecil terdiri dari sel
tunggal. Tubuh manusia minimal
mengandung 1014 sel. Terdapat
berbagi jenis sel yang amat bervariasi dalam ukuran, bentuk, dan fungsi
khususnya.
Dalam setiap organisme
multiseluler (misalnya manusia, tanaman
jagung dll.) terdapat puluhan atau ratusan jenis sel yang berbeda, semuanya
terancang secara khusus untuk bersama-sama berfungsi di dalam jaringan
dan organ. Namun demikian bagaimanapun
besar dan kompleksnya organisme tersebut, setiap jenis sel mempertahan kan sifat
khusus dan kebebasannya.
Meskipun berbeda dalam
fungsi dan penampilan, berbagai jenis sel menunjukkan kesamaan ciri struktur
dasar sebagai berikut :
1) Tiap sel dikelilingi oleh membrane tipis yang membuatnya
terpisah dan sampai tingkat tertentu mampu mencukupi diri sendiri. Membran sel yang juga disebut membrane
plasma atau membrane sitoplasma bersifat permeable selektif. Membran ini mengangkut nutrient dan garam
yang dibutuhkan ke dalam sel, dan produk buangan dari sel keluar. Membran ini biasanya tidak permeable terhadap
senyawa-senyawa yang ada di lingkungan sel yang tidak dibutuhkan. Pada semua sel susunan molekul membrane
plasma umumnya serupa yaitu terdiri dari dua lapis molekul lipid yang
mengandung protein khusus. Beberapa
protein membrane merupakan enzim, sementara yang lain dapat mengikat nutrient
dari lingkungan dan mengangkutnya ke dalam sel.
2) Di dalam tiap sel terdapat sitoplasma. Sitoplasma merupakan tempat berlangsung nya
hampir semua reaksi enzimatis dari metabolisme sel. Di sini sel menggunakan energi kimia untuk
membangun dan mempertahankan strukturnya, serta melakukan pergerakan sel atau
kontraksi. Di dalam sitoplasma sel juga
terdapat ribosom yaitu suatu
granula kecil berdiameter antara 18 – 22 nm yang berfungsi mensintesa protein.
3) Semua sel hidup dilengkapi dengan inti sel atau suatu badan serupa
inti. Inti sel merupakan tempat
terjadinya replikasi senyawa genetic dan penyimpanan dalam bentuk asam
deoksiribonukleat (DNA =
deoxyribonucleic acid).
2.4.
Struktur sel Hidup
Sel hidup senantiasa mengandung
protoplasma karena protoplasma didefinisikan sebagai isi sel hidup, dan tidak
mencakup dinding sel. Protoplasma sebuah sel disebut protoplas. Dengan
demikian, sel dapat dibagi menjadi (1) protoplas,
yakni seluruh bagian dalam sel, dan (2) dinding
sel yang mengelilinginya.
2.4.1.
Protoplas
Protoplas terdiri atas bagian
bermemberan dan tak bermemberan. Dengan memakai pengawet yang cocok, dan
disiapkan guna diamati dengan mikroskopelektron transmisi, penampang melintang
memberan tampak seperti dua garis hitam, masing-masing seteba l2.5 nm,
dipisahkan oleh garis pucat setebal 3.5 nm,sehingga disebut memberan ganda yang
dipisahkan oleh ruang sempit. Jenis memberan seperti itu disebut satuan memberan.
Pada umumnya dianggap bahwa satuan
memberan terdiri atas lapisan lipida yang disusun oleh lapisan dua molekul dan
dibatasi disetiap sisinya oleh selapis protein. Protein dalam satuan memberan
tersusun dalam pola khusus yang menyebabkan organel yang memiliki satuan
memberan seperti itu memiliki sifat permeabilitas khusus. Memberan merupakan
bagian sitoplasma yang hidup dan dapat berubah sesuai dengan aktivitas
sitoplasma. Satuan memberan bersifat permeabilitas
selektif terhadap berbagai zat yang melewatinya. Dewasa ini memberan berfungsi
menurut model mozaik cair diterima
secara umum.
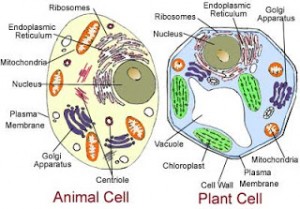
Protoplas dapat dibagi menjadi
sitoplasma dan nukleus. Sitoplasma meliputi reticulum endoplasma, diktosom,
mitokondria, plastisida, mikrobodi, ribosom,
sferosom, mikrotubul, mikrofilamen, vakuola, dan zat ergastik (gambar 2.1.).
Sitoplasma adalah bagian protoplasma berupa cairan kental atau yang lebih pekat
seperti agar-agar. Kedua bentuk tersebut tidak hanya berbeda dan bentuk yang
satu dapat dengan mudah berubah menjadi bentuk lainnya. Sebagian besar (85-90%)
sitoplasma terdiri dari air, disamping senyawa yang berada dalam larutan
sebagai koloid atau terlarut.
2.4.2. Susunan kimiawi
sitoplasma
Secara umum sitoplasma terdiri
dari oksigen, karbon, hydrogen, dan nitrogen. Oksigen berjumlah sekitar 62%,
karbon 20%, hidrogen 10 %, dan nitrogen 3%. Sisanya yang 5% terdiri dari
sekitar 30 unsur; yang terpenting di antaranya adalah Ca, Fe, Mg, Cl, P, K,dan
S. Selain itu, ditemukan juga dalam jumlah yang lebih kecil: Bo, Cu, Fl, Mn,
dan Si. Pada sel tertentu dapat ditemukan alcohol, Co, dan Zn. Semua unsur
tersebut terdapat dalam bentuk ion atau melekat pada molekul karbon, Fosfor,
misalnya terdapat dalam ATP (adenosine trifosfat). Di atas telah dissebutkan
bahwa sitoplasma terdiri dari airsampai 85-90%. Gas seperti Co2 dan
O2 tetap larut di dalamnya.
Protoplasma setiap sel mengandung
zat organic, dan yang penting di antaranya adalah karbonhidrat, lemak, dan
protein.
2.4.2.1. Karbohidrat
Sekitar 13%
protoplasma terdiridari karbohidrat, yang mengandung karbon, hidrogen, dan
oksigen. Karbohidrat yang terpenting adalah glukosa, sukrosa, pati selulosa.
Karbohidrat berada dalam bentuk suspensi atau terlarut dalam protoplasma, dan
tugas utamanya adalah menghasilkan energi.
2.4.2.2. Lemak
Lemak atau
lipida terdiri dari unsurkarbon, hidrogen, dan oksigen. Lemak dibentuk oleh
gliserol dan asam lemak. Dibandingkan dengan karbohidrat, jumlah oksigennya
kurang. Penguraian lemak secara kimiawi menghasilkan energi yang jumlahnya
lebih besar dibandingkan dengan yang dihasilkan karbohidrat.


Gambar 2.1. A,Bagian
tiga dimensi sel tumbuhan dan organel yang ada di dalamnya.B, bagan
dua dimensi sel tumbuhan yang meristematik. (A
dari Mauseth, 1988 dari Sitte,
1961, dalam Fahn, 1989)
2.4.2.3. Protein
Sekitar 15% protoplasma terdiri dari protein. Selain unsur karbon,
hidrogen, dan oksigen, protein terutama mengandung nitrogen, dan biasanya
terdapat pula P dan S. Protein dibentuk oleh asam amino. Ada 20 macam asam
amino di alam, yang membentuk berbagai macam protein dalam kombinasinya yang
berbeda-beda. Protein terlarutkan pleh ensim proteolitik. Sering kali protein
berkombinasi dengan zat lain, terutama dengan asam nukleat (nucleoprotein),
karbohidrat (glikoprotein), dan lemak (lipoprotein).
2.4.2.4. Nukleoprotein
Dalam protoplasma terdapat dua macam asam nukleat, yakni asam ribonukleat
(ARN) dan asam deoksiribonukleat (ADN). ARN terdiri dari sejumlah nukleotida
yang saling berkaitan membentuk rangkaian tunggal yang panjang. Sementara itu,
ADN terdiri dari sejumlah besar nukleotid yang terangkai menjadi satu benang
tunggal. Dua benang seperti itu dapat tersusun sejajar karena berkaitan pada
basanya yang berpasngan dengan cara tertentu, serta dapat pula melingkar dalam
bentuk heliks. ADN dan ARN mengendalikan kegiatan metabolisme dalam sel. DNA
terutama ditemukan dalam kromosom dan menurunkan sifat dari hamper semua mahluk
hidup.
Disamping berbagai senyawa diatas, terdapat zat lain, terutama yang
berhubungan dengan metabolisme sel. Beberapa diantaranya ditemukan didalam
semua sel, dan beberapa hanya pada sel khusus seperti berbagai piqmen, lateks,
alkaloid, vitamin, hormon, dan antibiotika.
2.4.3.
Sifat fisika sitoplasma
Protein dalam sitoplasma
terdapat dalam koloid. Protein tersebar sebagai butiran yang sangat halus, dan
setiap butiran terdiri dari beberapa molekul. Umumnya protoplasma terbentuk
lir-emulsi dengan protein tersebar dalam air. Banyak partikel mampu
menghantarkan air. Hal itu disebut hidrasi dan koloid yang menahan air disebut
terhidrasi. Koloid pada sitoplasma tetap terhidrasi, namun kadar hidrasinya
bergantung pada macam-macam kondisi.
Sitoplasma hidup
bersifat bening dan transparan; indeks bias dan kemampuannya menyerap cahaya
tidak terlalu berbeda sehingga dalam keadaan hidup tak terlihat nyata. Dalam
arti luas, istilah sitoplasma dipakai bagi zat protoplasma yang mengelilingi
inti dan organel lain. Istilah seperti sitoplasma dasar dan sitosol dipakai
untuk bagian sitoplasma di luar organel yang dibatasi memberan.
2.5.
Struktur organel
Dalam sitosol terdapat
sitoskeleton yang mendukung bentuk sel dan mampu menempatkan organel serta
menggerakannya. Sitoskeleton tersusun oleh jalinan filament protein, yang
paling penting di antaranya adalah mikrofilamen aktin dan mikrotubul. Didekat
dinding sel, sitoplasma dibatasi oleh memberan plasma yang disebut plasmalema,
sedangkan yang berbatasan dengan vakola disebut tonoplas. Bagian sitoplasma di
antara kedua memberan tersebut berbutir, dan sering disebut polioplasma. Dengan
mikroskop cahaya, batas-batas tersebut tak selalu dapat diperlihatkan,namun
bukti fisiologi senantiasa ada.
2.5.1. Retikulum endoplasma
(RE)
Dalam sitoplasma terlihat jalinan struktur
memberan (gambar 2.1). Memberan itu dibangun oleh lipid dan protein serta
membentuk system pipa halus yang beranastomosis. System tersebut disebut
disebut reticulum endoplasma (RE) yang tersusun dari satuan memberan ganda
sebagaimana diterangkan terdahulu. RE dapat berbentuk sisterna (wadah, tempat)
yang melebar, tubul (bentuk pipa) atau lapisan terlubang. RE juga berada dalam
plasmodesmata, yaitu benang sitoplasma yang menembus dinding bersama dari dua
sel yang berimpitan. Bagian RE yang terdapat di tengah sitoplasma berbentuk
tubula, dinamakan desmotubul (gambar 2.1).
Jika ribosom melekat pada
RE, maka RE disebut RE kasar atau RE berbutir. Bila tak ada ribosom, disebut RE
licin.adanya hubungan dengan ribosom diartikan bahwa RE terlibat dalm sintesis
protein. Pada jaringansekresi, RE licin diduga turut dalam produjsi senyawa
lipofil. RE dianggap turut berfungsi dalam angkutan dalam sel yaitu mengangkut
bahan yang disekresikan. Sisterna RE dapat melebar dan manghimpun protein dan
senyawa lain.
Diduga bahwa cara
melebarkan diri, sisterna atau vesikula (kantung kecil) pada RE membentuk
vakuola.
2.3.2. Diktiosom
Diktiosom terdiri dari
tumpukan kecil vesikula pipih yang tepi-tepinya tidak rata, melainkan terbagi
menjadi tubul yang saling berhubungan menyerupai jala (gambar 3.3). Diktiosom
berfungsi dalam memeroses bahan ketika digerakan melalui sel atau ke luar sel. Beberapa
vesikula yang berasal dari reticulum endoplasma (RE) berkumpul di satu sisi
diktiosom (muka pembentuk), kemudian menyatu membentuk vesikula baru bagi
diktiosom. Sewaktu bahan dalam vesikula ini diproses, terutama dengan
menambahkan gula kepada protein, vesikula lain akan dibentuk di sisinya. Secara
serempak vesikula yang telah besar dan matang akan dilepas di sisi yang
berhadapan (muka pendewasaan), dan bergerak menjauh sambil membawa bahan yang
telah diproses.
Vesikula berada dalam diktiosom
hanya untuk sementara waktu saja sewaktu bergerak dari muka pembentuk sampai
muka pendewasaan. Jadi seakan-akan terjadi pola arus vesikula yang bergerak
dari RE ke organel lain. Selain itu, bahan yang memasuki diktiosom dapat
bergerak ke jalinan tubul di tepi. Tubul kemudian membengkak, melepaskan diri,
dan bergerak menjauh.
Fungsi utama vesikula yang
diproses dalam diktiosom adalah dalam sekresi karbohidrat, misalnya nektar,
bahan dinding sel, lender, atau ikatan antara gula dan protein, yakni
glikoprotein dalam beberapa macam lender dan dinding. Jumlah diktiosom dalam
sel tumbuhan kurang dan lebih sedikit dibandingkan dengan pada sel hewan karena
tumbuhan kurang bersekresi. Pada tumbuhan, diktiosom dalam sel biasanya tidak
saling berhubungan atau sedikit saja hubungannya. Pada sel hewan, diktiosom
seringkali bergerombol dan saling berhubungan membentuk system atau benda besar
yang dinamakan Badan Golgi.
2.3.3. Mitokondria
Mitokondria merupakan
organel yang dapat dilihat dengan mikroskop cahaya jika sel hidup diwarnai
dengan Janus Green. Dalam preparat untuk mikroskop electron, mitokondria
terlihat dalam macam-macam bentuk seperti bulat, memanjang, dan kadang-kadang
bercuping.
Namun, dengan teknik yang
membangun kembali organel itu dari sayatan seri ditemukan bahwa ada pula yang
berbentuk silinder panjang yang bercabang. Panjang mitokondria mencapai 3 µm,
dengan garis tengah 0,5 – 1 µm, serta dibatasi oleh selubung yang terdiri dari
dua satuan memberan. Satuan memberan sebelah dalam membentuk tonjolan seperti
lipatan (krista) ke arah stroma (bahan dasar dalam mitokondria).
Stroma terdiri terutama dari protein. Dalam mitokondria ditemukan ribosom
yang lebih kecil ukurannya daripada yang terdapat dalam sitoplasma. Rribosom
itu mengandung fibril ADN, namun kemampuan genetiknya terbatas. Mitokondria
berfungsi dalam respirasi aerob dan memasok ATP sebagai sumber energi utama
bagi sel. Selain itu, mitokondria terlibat pula dalam metabolisme zat antara.
Pembentukan mitokondria mungkin sekali terjadi dengan cara pembelahan.
2.6.
Plastida
Plastida merupakan
organel yang amat dinamis dan mampu membelah, tumbuh, dan berdiferensiasi
menjadi berbagai bentuk. Pada sel muda tumbuhan tinggi, plastida biasanya tak
berwarna dan disebut leukoplas atau proplastida.
Pada daun, plastida berwarna hijau dan disebut kloroplas, serta
pada buah masak kadang-kadang kuning atau merah, disebut kromoplas. Pada sel
yang tidak menjadi hijau, seperti sel epidermis atau sel rambut tangkai sari
(misalnya pada Rhoeo discolor), plastida tetap tak berwarna, disebut leukoplas
(dalam arti sempit).
Leukoplas juga terdapat
pada jaringan yang tak terdedah pada cahaya. Pada jaringan semacam ini seperti
pada umbi, leukoplas membentuk butir pati yang disebut amilopas. Sataloit
adalah amilopas khusus dalam tundung akar dan pada buku beberapa batang muda,
serta terlibat dalam gaya berat. Leukoplas membentuk minyak atau lemak, dan disebut elaiopas, misalnya
pada epidermis daun Vanilla.
2.6.1. Kloroplas
Kloroplas pada umumnya berbentuk seperti lensa, biasanya berukuran 4 – 6
µm. Didalam kloroplas terdapat zat hijau daun atau klorofil, dan sedikitnya dua
zat warna kuning atau merah, atau kelompok zat warna (karotenoid): satu macam
karoten atau lebih (C40H56) dan xantofil (C40H56
O2). Kloroplas berfungsi dalam fotosintesis dan pada
kebanyakan tumbuhan berfungsi pula dalam
pembentukan pati dari karbohidrat terlarut hasil fotosintesis, serta melarutkan
kembali.
2.6.2. Kromoplas
Warna kuning, merah, atau merah bata pada kromoplas disebabkan oleh
kandungan karotenoidnya. Kromoplas sering kali berasal dari kloroplas, namun
dapat pula berasal dari proplastida. Yang penting dalam diferensiasi kromoplas
adalah sintesis dan penempatan pigmen karotenoid seperti karotenoid (pada
wortol, Daucus) atau likopen (pada tomat, Lycopersicon). Perkembangan pigmen
berkaitan dengan modifikasi, bahkan perombakan sama sekali, tilakoid. Dalam
peroses itu, globula (gelembung) lipid bertambah banyak. Dalam beberapa
kloroplas, pigmen disimpan dalam globula (cabe kuning, jeruk). Pada kromoplas
lain, pigmen berkumpul dalam fibril protein yang berjumlah banyak (cabe merah).
Bentuk ketiga dari pigmen adalah bentuk kristaloid. Pada tomat merah, perkembangan likopen
berbentuk Kristal berkaitan dengan memberan tilakoid. Beberapa Kristal menjadi
amat panjang dan tilakoid memanjang, sementara likopera dibentuk. Kristaloid
karoten dalam akar wortel dibentuk sewaktu struktur dalam plastida rusak dan
tetap berhubungan denganselubung lipoprotein (gambar 3.5) Kromoplas tidak
memiliki klorofil. Kromoplas sering berasal dari kloroplas, seperti pada kulit
buah jeruk yang berubah dari hijau menjadi merah kuning. Keadaan sebaliknya
dapat pula terjadi, seperti kromoplas pada akar wortel yang terbukti mampu
berdifrensiasi menjadi kloroplas. Pigmen karoten hilang dan tilakoid yang
membentuk klorofil dapat berkembang dalam plastida.
Kromoplas memberi warna
pada berbagai bagian alat tumbuhan. Namun, tidak seluruh warna pada tumbuhan
disebabkan oleh pigmen dalam plastida, sebab dalam cairan vakuola juga dapat
ditemukanberbagai zat warna.
2.6.3. Amiloplas
Di beberapa tempat
tertentu, kloroplas membentuk butir pati besar sebagai cadangan makanan, seperti pada umbi semu
anggrek. Namun, jumlah cadangan makanan terbesar dibentuk dalam leukoplas umbi
baker, umbi batang, rizoma, dan biji.
Amilum atau pati dapat ditunjukkan dengan mudah karena berwarna
biru atau hitam dengan idodium. Bila dipanaskan sampai 70oC warna
hilang dan menjadi biru lagi setelah dingin kembali. Reaksi ini dianggap
sebagai permukaan. Butir besar menunjukkan lapisan yang mengelilingi sebuah
titik di tengah, yakni hilum. Hilum biasa berada di tengah butir pati agak ke
tepi. Retakan yang sering terlihat berarah radial dari hilum nampaknya terjadi
akibat dehidrasi butir pati.
Terjadinya lapisan, dan
secara bertahap menjadi lebih renggang di sebelah luar.dianggap menyebabkan
perbedaan kadar air yang terkandung di dalamnya. Jadi, adanya lapisan dianggap
akibat perbedaan kadar air dalam lapisan
yang berturut-turut, sedangkan taraf kepadatan menyebabkan perbedaan indeks bias.
Dalam alcohol kuat, semua
lapisan itu hilang, mungkin karena dehidrasi yang meniadakan perbedaan
taraf kepadatan. Pada pati serealia,
terjadinya lapisan bergantung pada irama harian.
Pada Kentang ,
perubahan berkala yang mengakibatkan
adanya lapisaan berasal dari dalam
(endogen). Dalam butir tersebut. Pada biji yang mulai berkecambah atau umbi
yang mulai menumbuhkan pucuk, butiranti mengalami pengikisan yang bermula dari
luar dan lama – kelamaan habis terurai. Pada butir pati kecil, hilum biasanya menjadi eksentris (tidak di pusat). Jika
dalam plastida terbentuk lebih dari satu butir pati, maka butiran tersebut akan
segera saling menyentuh dan membentuk butir majemuk.
Dengan demikian dikenal butir majemuk seperti pada pati
gandum (Avena) dan padi (Oryzasativa), pati setengah majemuk
pada kentang, dan butir pati tunggal seperti pada pati irut (Maranta).
Jika butir pati mengisi sel
hingga penuh, maka tepinya bersudut. Posisi hilum, bentuk dan ukuran
butir, serta sifat butir tunggal atau majemuk memungkinkan identisifikasi
spesies tumbuhan penghasilan butir pati yang bersangkutan.
2.7. Struktur dan perkembangan kloroplas
Semua jenis plastida
berasal dari butiran proplastida yang terdapat dalam sel meristem dan sel
telur. Plastida atau proplastida memperbanyak diri dengan membelah. Dalam
berbagai plastida, termasuk kloroplas, bias ditemukan lipid berbentuk globul
(Plastoglobuli) dan fitoferitin (senyawa besi-protein). Selain itu, dilaporkan
adanya tubuh protein amorf yang dibatasi oleh membrane. Sistem membrane pada
plastida disusun oleh sejumlah kantung
yang dinamakan tilakoid.
Pada proplastid, system
tilakoid merupakan yang paling rendah
taraf perkembanga uannya dan terdiri hanya dari beberapa tilakoid atau
tak ditemukan tilakoid sama sekali. Di saat plastida berdeferensiasi, membrane
dalam menguncup membentuk vesikula pipih, kemudian berproliferasi (tumbuh
meluas) dan membentuk tilakoid. Dalam kloroplas, tilakoid menempatkan dirinya
menurut susunan yang khas. Pada saat kloroplas berdiferensasi, jumlah
ribosom meningkat, sedangkan pada
leukoplas, produksi ribosom lengkap terhenti dan jumlahnya berkurang sejak wal
stadium perkembangannya.
Dalam kloroplas, system
tilakoid terdiri dari grana dan fren (tilakoid dalam stroma). Setiap grana terdiri dari satu tumpukan
tilakoit yang masing-masing berbentuk cakram. Grana saling berhubungan
dengan adanya jalinan fret dalam stroma.
Kini dianggap bahwa kedua jenis tilakoid
saling berhubungan sehingga ruang di dalamnya bersinambungan. Membran kloroplas
terdiri dari lipid dan protein dalam jumlah yang sama.
Klorofil terdapat pada membrane tilakoid. Pada
membrane itu banyak terdapat pada
membrane tilakoid. Pada membrane itu banyak terdapatpartikel persatuan. Jenis
partikel tertentu dianggap sebagai bagian dari membrane yang namakan kuantosom,
yakni satuan morfologi untuk berlangsungnya reaksi fotosinteis yang memerlukan
cahaya. Namun, tampaknya kuantosom tidak perlu selalu berbentuk satu unit
fungsi. Selain system untuk menangkap cahaya matahari, dalam kloroplas juga
terdapat enzim yang membantu fiksasi CO2 menjadi gula.
Pada tumbuhan tinggi yang
tumbuh di tempat gelap,daun serta batang yang berkembang tampak pucat, disebut,
teretiolasi. Dalam ikeadaan seperti itu,
vesikula yang berasal dari membran dalam berkembang menjadi kerangka
parakristal yang disebut tubuh prolamela. Plaspetid seperti itu dinamakan
etioplas. Jika kemudian tumbuhan disimpan di tempat bercahaya. Maka tubuh
prolamela berkembang menjadi system tilakoid yang khas bagi kloroplas.
Sebagaimana
diutarakan di atas, baik plastida maupun mitokondra mengandung ADN dan
ribosom. Sebab itu kedua organel tersebut memiliki kemungkinan untuk
berotonomi. Indikasi seperti itu mengakibatkan adanya hipotesis yang menyatakan
bahwa dalam perkembangan evolusi, plasida dan mitokondria berasal dari
prokariot (misalnya, sejenis alga biru) yang terkandung dalam sel eukariot yang
primitive dan di sana memantapkan diri sebagai tubuh simbiotik.
2.8. Mikrobodi
Nama “mikrobodi” diberikan kepada
tubuh kecil-kecil dalam sel yang semula
tak diketahui fungsinya. Namun, kini diketahui bahwa sedikitnya terdapat dua
kelompak mikrobodi : (1) peroksisom yang berhubungan dengan kloroplas dan merupakan
tempat fotorerespirasiasamglikolat,dan(2)glioksisom yang terlibat dalam
penggunaan asetil-CoA pada waktu pengerahan cadangan lipid, terutama sewaktu
perkecambahan biji yang mengandungminyak.
2.9. Ribosom
Ribosom
merupakan partikel kecil bergaris tengah 17 - 20 !-lm, berfungsi dalam sintesis
protein. Ribosom terdiri dari dua bagian yang masing-masing tersusun oleh ARN
ribosom (rRNA) dan protein. Kedua bagian tersebut disintesis di nukleolus,
bergerak ke luar inti ke sitoplasma, kemudian berikatan sesamanya dan dengan
ARN duta (mRNA) untuk mentranslasikannya ke dalam protein. Karena ARN duta
cukup besar dan panjang dan dapat dibaca sekaligus oleh beberapa ribosom
berikut translasinya, maka ribosom sering ditemukan dalam kelompok. Kelompok
itu dinamakan polisom atau poliribosom
Polisom
ditemukan bebas dalam hialoplasma atau melekat pada RE sehingga disebut RE
kasar. ARN Transfer (tRNA) terdapat dalam semua sel, namun paling banyak
ditemukan dalam sel dengan kecepatan sintesis protein paling tinggi, seperti
sel penyimpan protein dalam beberapa macam biji.
Dalam
plastida dan mitokondria terdapat ribosom 70S yang lebih mirip dengan yang
ditemukan pada prokariot. Ribosom yang disintesis dalam inti dan yang berfungsi
dalam sitoplasma agak berbeda dalam ukuran, bentuk, serta kepekaan terhadap
antibiotika dinamakan ribosom 80S.
2.10. Sferosom
Sferosom
adalah tubuh lipid berbentuk bulat, yang dalam mikroskop tampak buram setelah
diawetkan dengan osmium tetroksida. Beberapa peneliti berpendapat bahwa
sferosom dibatasi oleh membran, sedangkan peneliti lain menganggap bahwa batas
luar organel ini terdiri dari selapis molekul lipid yang dibentuk sebagai
tanggapan terhadap sitoplasma sekelilingnya yang mengandung air.
2.11. Mikrotubul
Mikrotubul
terdiri dari tubul protein yang lurus dan ramping (garis tengah 23 - 27 !-lm).
Mikrotubul amat penting karena terlibat dalam membentuk serta mempertahankan
bentuk sel serta diferensiasi kimiawi. Pada sel hewan, mikrotubul dianggap
menjadi bagian dari kerangka (skelet) sel, namun pada tumbuhan mungkin lebih
sesuai disebut templat atau pagar/batas. Mikrotubul menghasilkan bentuk
sementara yang kemudian dipertahankan secara permanen oleh dinding. Susunan
mikrotubul dapat menangkap dan memandu vesikula ke tempat sintesis atau dapat
memisahkannya dari tempat seperti itu. Telah ada bukti yang menunjukkan bahwa
mikro tubul mengarahkan fibril selulosa di saat dinding dibentuk.
Pada
waktu mitosis, kumparan dibentuk oleh mikrotubul yang mengarahkan dan
menggerakkan kromosom. Sesudah itu, sewaktu sitokinesis, mikrotubul membentuk
fragmoplas yang mengendalikan peletakan, penempatan, dan dinding baru.
2.12. Dinding sel
Adanya dinding membedakan sel
tumbuhan dari sel hewan. Dinding sel telah banyak diteliti karena kepentingannya
dari segi biologi maupun komersial. Informasi itu ditunjang oleh penelitian
dari segi kimia, biokimia, fisika, dan morfologi. Dinding sel dihasilkan
protoplas ke arah luar. Senyawa yang terutama terdapat di dalamnya adalah
selulosa. Senyawa lain adalah hemiselulosa, pektin, protein, serta zat seperti
lignin (zat kayu), dan suberin (zat gabus).
2.12.1. Struktur dinding
sel
Berdasarkan perkembangan dan
strukturnya dikenal tiga lapisan dinding, yakni
lamela
tengah, dinding primer, dan dinding sekonder. Lamela tengah adalah
bagian yang melekatkan kedua sel yang berdampingan dan berada di an tara
dinding primer kedua sel tersebut. Lamela tengah terdiri terutama dari pektin.
Enzim pektinase dapat melarut lamela tengah sehingga sel akan terpisah satu
sarna lain. Proses itu disebut maserasi.
Dinding primer adalah dinding yang dibentuk ketika sel
sedang tumbuh. Dinding ini terdiri terutama dari selulosa yang disertai zat
bukan selulosa sepert hemiselulosa dan senyawa berpektin, serta bersifat optis
aktif (anisotrop). Dinding sekunder terbentuk di sebelah dalam dinding
primer, setelah sel selesai tumbuh. Lapisan dinding ini berkerangka selulosa
sebagai unsur utama, disertai berbagai zat nonselulosa, namun biasanya tak
mengandung senyawa berpektin. Lignin (zat kayu) bisa ditemukan. Jika tetjadi
lignifikasi, maka lignin mula-mula terdapat di lamela tengah, kemudian di
dinding primer, dan akhirnya di dinding sekunder.
Molekul selulosa dalam dinding
berhimpun menjadi sejumlah berkas yang disebut mikrofibril. Di beberapa
bagian mikrofibril, susunan molekul selulosa sangat teratur. Bagian seperti itu
disebut mise!. Ruang yang berada di antara mikrifibril dapat terisi
dengan senyawa lain seperti lignin dan suberin (zat gabus). Sejumlah berkas
mikrofibril membentuk makrofibril.
Sinar polarisasi menunjukkan bahwa
selulosa bersifat kristal, molekulnya tersusun amat teratur. Sifat kristal
dapat diperlihatkan secara sederhana dengan cara menempatkan selembar polkaroid
di antara sumber cahaya dan mikroskop, dan selembar polaroid lain di antara
lensa okuler dan mata. Cahaya yang melalui suatu polaroid akan bergetar dalan
satu bidang saja-cahaya terpolarisasi dalam satu bidang. Jika kedua polaroid
tersebut dipasang tegak lurns sesamanya (bersilangan) dari segi bidang
polarisasinya, maka lantang pandangan dalam mikroskop akan tetap gelap.
Jika suatu zat bersifat kristal
seperti selulosa ditempatkan di antara kedua seluloid yang bersilang itu, maka
hanya sedikit cahaya yang bisa dilewatkan dan mencapai mata. Disebabkan oleh
sifat kristal dan sifatnya yang membias ganda, maka selu!osa mengubah bidang
polarisasi cahaya yang mencapai polaroid di atas okuler dan sebab itu
mengakibatkan cahaya mencapai mata. Pada posisi polaroig tertentu, pembiasan
ganda tidak diperlihatkan. Senyawa yang mempengaruhi cahaya seperti halnya
selulosa disebut bersifat anisotrop. Senyawa yang tidak mempengaruhi cahaya
disebut bersifat optis tidak aktif atau isotrop.
Senyawa pektin dianggap isotrop
oleh sebab itu lamela tengah tampak sebagai
bagian hitam jika diamati dengan mikroskop polarisasi dengan polaroid
dalam posisi silang. Sebaliknya, dinding primer dan dinding sekunder adalah
anisotrop karena kandungan selulosanya tinggi.
Cara dinding sel tumbuh, jumlah
perturnbuhan, serta susunan rnikrofibril dalarn penarnbahan senyawa selanjutnya
dalarn dinding sel rnengakibatkan terjadinya lapisan dinding sel. Lapisan
dinding tertua adalah yang terluar, sedangkan bagian termuda adalah yang
terdalarn, yakni yang berbatasan dengan protoplas.
Dalam sel kayu yang berdinding
tebal sering dapat dibedakan 3 lapisan utama yang diberi nama SI, S2, dan S3.
Lapisan S2 paling tebal. Kadang-kadang S3 cukup berbeda dari yang lain sehingga
disebut lapisan dinding tersier. Perbedaan lapisan-lapisan sering tak
jelas dan lamela tengah beserta dinding primer di kedua sisinya sering tampak
sebagai satu lapisan saja dan disebut lamela- tengah majemuk.
Pemisahan dinding sekunder menjadi
tiga lapisan S diakibatkan terutama oleh letak mikrofibril dalam ketiga lapisan
itu. Pada umumnya mikrofibril dalam dinding cenderung berbentuk sebagai heliks.
Dalam SI kemiringan heliks membentuk sudut besar dengan sumbu panjang sel sehingga
letak mikrofibril hampir datar (gambar 3.9). Pada S2, sudutnya kecil, dan
kemiringannya terjal. Dalam S3, mikrafibril tersusun seperti pada Sl, bersudut
besar dengan sumbu panjang sel. Dinding primer berbeda dari dinding sekunder
dalam susunan mikrofibrilnya yang acak.
Dalam sel yang memiliki dinding
sekunder, dinding primer biasanya am at tipis. Hal itu juga terlihat pad a sel
parenkim yang aktif dalam metabolisme seperti mesofil daun dan parenkim
menyimpan cadangan makanan dalam umbi-umbian. Namun, dinding primer menjadi
amat tebal pada sel kolenkim di batang dan daun serta endosperm biji.
2.12.2. Susunan kimiawi
Senyawa penting yang terutama dalam dinding sel
adalah selulosa, kemudian hemiselulosa, senyawa pektin, dan protein. Selulosa,
suatu polisakarida, adalah polimer yang tersusun oleh molekul glukosa dalam
ikatan beta 1,4 dan membentuk rantai IUTUS yang panjangnya mencapai 4
mikrometer. Rumus empirisnya (CuHlOOs)n.
Mo1ekul selulosa mengandung 8.000
- 15.000 monomer glukosa dan mencapai panjang 0,25 - 5,0 !-lm. Sifat mekanik
selulosa yang paling utama adalah kekuatan rentangnya. Pada tekanan yang
menempatkan mikrofibril, selulosa akan melengkung. Hal ini dihindari dengan
mengganti matriks antarfibril selulosa dengan zat padat yang membuatnya menjadi
keras. Proses pemasukan zat tambahan ke dalam dinding sel dengan menempatkannya
dalam matriks antarfibril selulosa dinamakan pelapisan atau pengerasan.
Pengerasan paling penting pada
tumbuhan tinggi adalah penempatan ligin, suatu polimer yang tersusun oleh
satuan fenilprapanoid. Lignin menyebabkan dinding menjadi keras dan kaku.
Selain itu, suberin (zat gabus), kutin, malam, tanin, dan zat lain dapat
menempatkan diri di dalam matriks. Di samping itu, selulosa bergabung dengan
berbagai polisakarida lain seperti hemiselulosa dan zat pektin (senyawa
poliuronida). Air pun terdapat dalam dinding dalam jumlah yang berbeda-beda,
sesuai dengan sifat sel yang bersangkutan. Beberapa macam hemiselulosa
nampaknya berperan sebagai penghubung penting antara polimer non-selulosa dan
selulosa.
Kerangka selulosa tersusun oleh
sistem fibril (serabut) yang terdiri atas molekul selulosa. Molekul selulosa
mengandung 8.000 - 15.000 monometer glukosa dan mencapai panjang 0,25 - 5,0
mikrometer trap.
Setiap mikrofibril mengandung 40 -
70 rantai selulosa yang sejajar letaknya. Kelompok enzim pensintesis selulosa
tertanam pada plasmalema dalam bent uk roset . Enzim itu dianggap menerima
glukosa yang teraktivasi dari sisi sitoplasma dan membubuhkannya kepada molekul
selulosa yang sedang tumbuh yang muneul di sisi plasmalema yang menghadap
dinding. Karena enzim pensintesis selulosa terkumpul dalam roset, maka molekul
yang tumbuh dengan sendirinya berada di tempat yang teratur dan segera
mengkristal.
Dalam
sistem seperti itu dapat diperkirakan bahwa mikrofibril yang tumbuh akan
berinteraksi dengan bahan dinding sebelumnya dan akan terpaneang dengan kukuh.
Jadi, sewaktu pertumbuhan molekul berlanjut, kumpulan enzim yang berbentuk
roset itu dapat bergerak maju di bidang plasmalema. Hal ini dapa~ dipahami jika
dianut anggapan bahwa struktur membran mengikuti model mozaik cairo
Roset, yang
berupa kumpulan enzim pensintesis selulosa, dapat beragregasi menjadi sampai 16
baris. Keseluruhannya bergerak melalui membran sebagai satu kesatuan dan
menghasilkan baris-baris kristal selulosa yang sejajar. Gerakan struktur itu
amat khas sehingga baris-baris roset tersusun bersama dan mikrofibril baris
satu dapat bergabung dengan mikrofibril baris yang lain menghasilkan struktur
1ebih besar yang disebut makrofibril. Makin banyak roset dalam satu
baris, makin banyak jumlah mikrofibril dalam satu makrofibril, dan makin besar
pula garis tengahnya. Makrofibril
seperti itu lebih khas bagi dinding sekunder yang terbentuk setelah sel
berhenti tumbuh.
Dengan
metode penambahan seperti di atas, mikrofibril atau fibril baru diletakkan di
bagian dalam unsur dinding sebelumnya dan menghasilkan struktur dinding yang
berlapis-lapis. Proses penambahan bahan dinding baru yang hanya ke arah dalam
disebut aposisi. Mikrofibril dapat pula tumbuh menghasilkan jalinan
dengan adanya mikrofibril yang tumbuh di antara mikrofibril lama. Proses itu
sering dinyatakan dengan istilah intususepsi.
Ada dua
teori yang menerangkan cara pertumbuhan dinding primer. Yang pertama disebut teori
mozaik yang menjelaskan bahwa tekstur serabut dalam bagian dinding sel
tertentu merengang akibat tekanan turgor dan kemudian menjadi padat kembali
dengan peletakan mikrofibril baru dalam celah yang disebabkan tekanan tersebut.
Perenggangan jalinan fibril memerlukan kekenyalan (keliatan) matriks dinding
pada daerah yang meluas. Zat pengatur tubuh dan enzim yang terdapat dalam dinding
terlibat dalam pengaturan penambahan luas ini.
Teori
kedua adalah pertumhuhan multinet. Menurut konsep ini, penebalan
danpenambahan luas di~din~ priI?e~ sering diakibatkan aleh pemisaha_n_
mikrofibril yang bersilangan dan perubahan arahnya pada lamela yang dibentuk
paling awal, dari hampir datar - .. menjadi hampir tegak. Lamela baru yang
disusun oleh mikrofibril lebih padat, bersilangan, dan berarah hampir datar;
ditambah secara sentripetal.
Selain
berperan dalam sintesis molekul selulosa enzim pada dinding sel dapat pula
berperan dalam pemindahan (transfer) dan hidrolisisnya. Enzim itu juga berperan
dalam modifikasi metabolit dari luar sel yang memudahkan pengangkutannya ke
dalam sel. Ada pula enzim yang berperan sebagai protein struktur yang kaya akan
serin (suatu asam amino) dan dapat mengandung sampai 25% hidroksiprolin.
Fungsi
protein struktur pada dinding belum diketahui benar. Namun, karena cenderung
berlaku seperti batang panjang dan kaku, ada kemungkinan protein dapat mengikat
silang bagian lain dan dapat berlaku sebagai unsur struktur. Protein yang
berikatan dengan polisakarida ~ada dinding sel dianggap membentuk glikoprotein.
Penambahan senyawa tersebut
dilaporkan berkaitan dengan peristiwa meredanya pembentangan dinding sel.
Mekanisme apakah yang
memandu arah mikrofibril dan makrofibril? Penelitian menyatakan bahwa
mikrotubul menyusun suatu pola tepat di sebelah dalam plasmalema. Roset yang
terdapat dalam membran plasma dapat berinteraksi dengan mikrotubul dan mungkin
dipandu olehnya.
Selain selulosa, dalam dinding ban
yak terdapat hemiselulosa. Zat ini berupa campuran dari polimer yang berbeda
susunannya dan bercabang banyak. Hemiselulosa mempunyai punggung dasar
berikatan beta 1,4 yang bercabang-cabang ke arah lateral. Karena cabang lateral
dapat berinteraksi dengan selulosa, maka hemiselulosa mengkristal bersama
selulosa, melapisinya, dan melekatkan semua mikrofibril menjadi satu.